



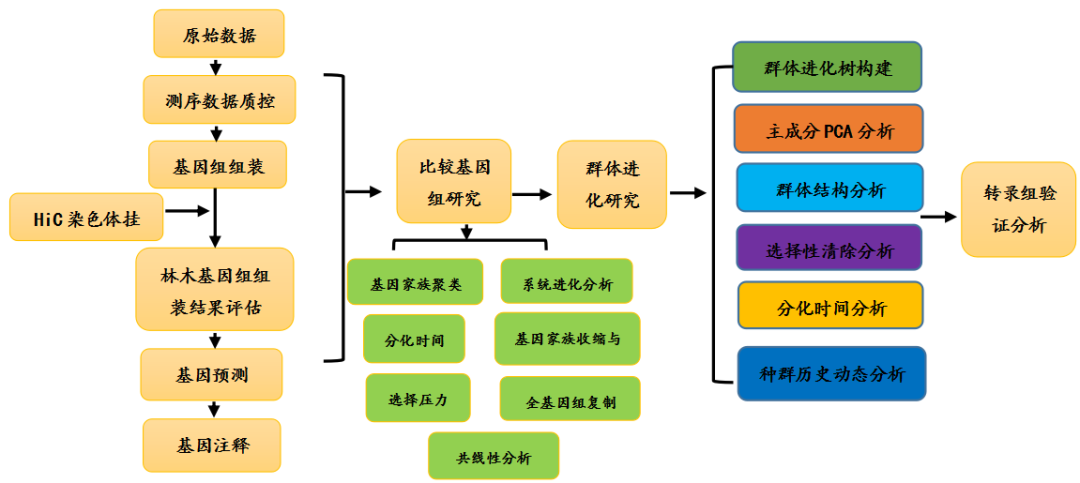
林木基因组研究技术路线
林木基因组重点研究思路(一)基因组+群体进化
案例(1):开心果基因组

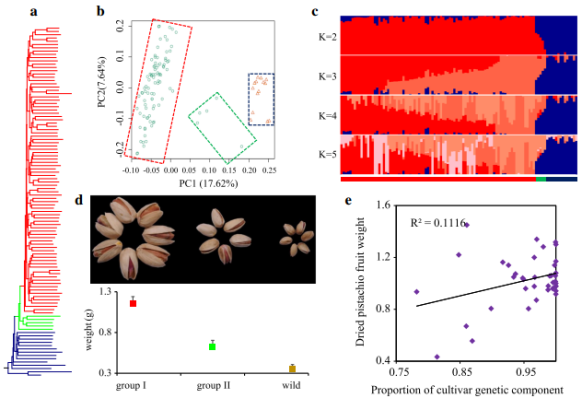
图1 体进化分析
研究结果1:揭示了开心果的两步驯化过程(栽培种质分为两个群。栽培种Group I的LD值最高,栽培种Group II和野生开心果的LD衰减值相近。Group II包括 Qazvini,Italiaei和Badami Zarand在内的5种类型的个体,且这些种质被记录为古代具有种子的材料);
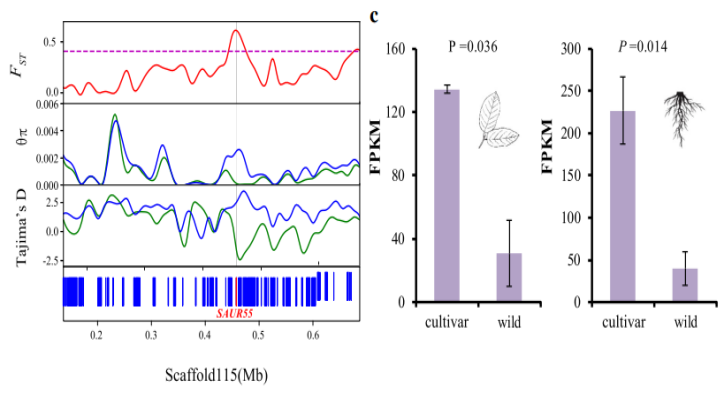
图2 开心果株型群体驯化(野生/驯化)分析
研究结果2:在开心果中,基因组上约有 9.2 Mb 的区域被鉴定为具有高水平的群体分化,包含665个基因,其中开心果株型可能与 SAUR55 基因的选择相关,同时该基因 表现出了显著增加的表达水平;果实重量的变化发生在驯化与人工选择期间(CYCD7-1基因)。
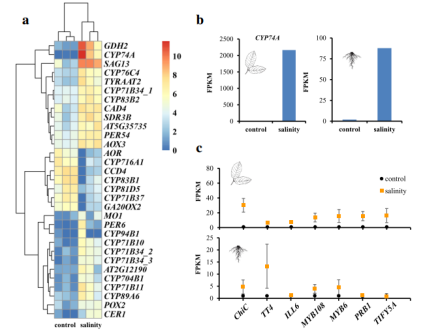
图3 开心果盐胁迫下转录组分析
研究结果:转录组研究分析发现编码几丁质酶的基因和参与 JA 生物合成途径的基因可能有助于开心果适应盐水环境。
案例(2):白桑基因组

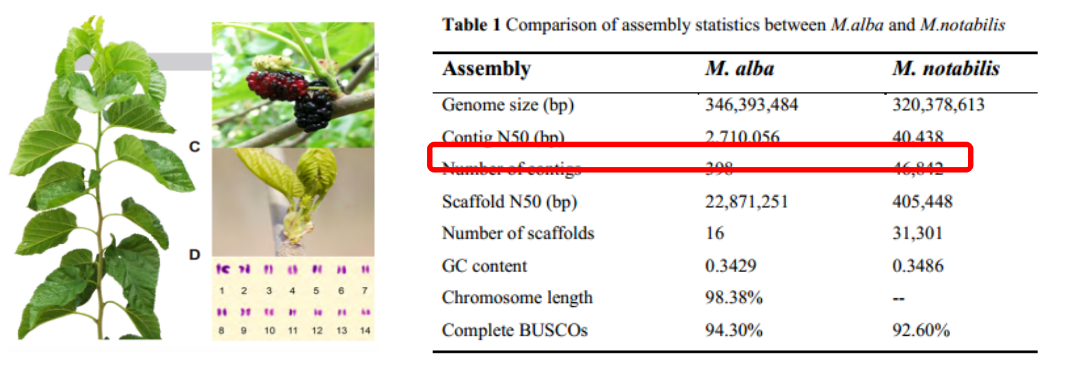
图1 白桑基因组组装结果
群体进化研究:群体材料选择(重测序:~15.6X):134份桑树种质(132份栽培种,来自中国和日本;2份野生种,来自中国陕西);
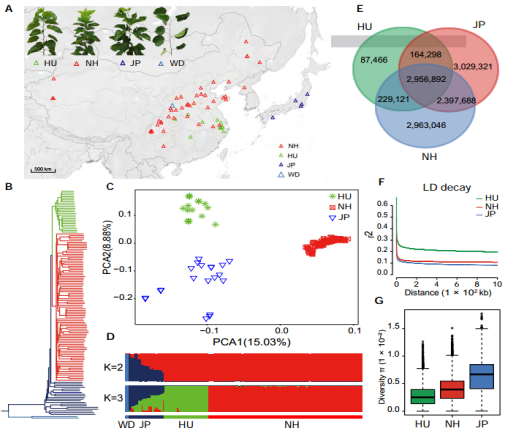
图2 桑树群体进化分析
研究结果1:利用得到的14 Mb SNP对广东桑以外的其余四种桑树进行了聚类分析,未得到与形态分类相似的聚类结果,基因组数据支持《中国植物志》中将白桑、鲁桑、山桑和瑞穗桑这4种栽培桑树种合并的结论。首次用基因组数据明确了栽培桑树物种分类,认为白桑、鲁桑、山桑和瑞穗桑都属于一个物种,即白桑(Morus alba?L)。
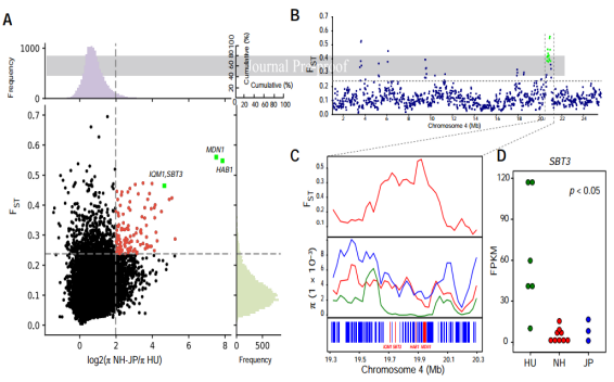
图3 湖桑(NH-JP/π HU)驯化群体选择研究
?研究结果2:湖桑祖先群体比非湖桑的祖先群体小,表明湖桑在太湖流域养蚕业的扩张过程中可能经历了强大的瓶颈或密集的人工选择;湖桑群体选择分析揭示驯化过程适应性机制,发现与植物生长,防御反应,茉莉酸途径相关基因受到选择(MDN1,HAB1,IQM1,SBT3)。
案例(3):小垫柳基因组

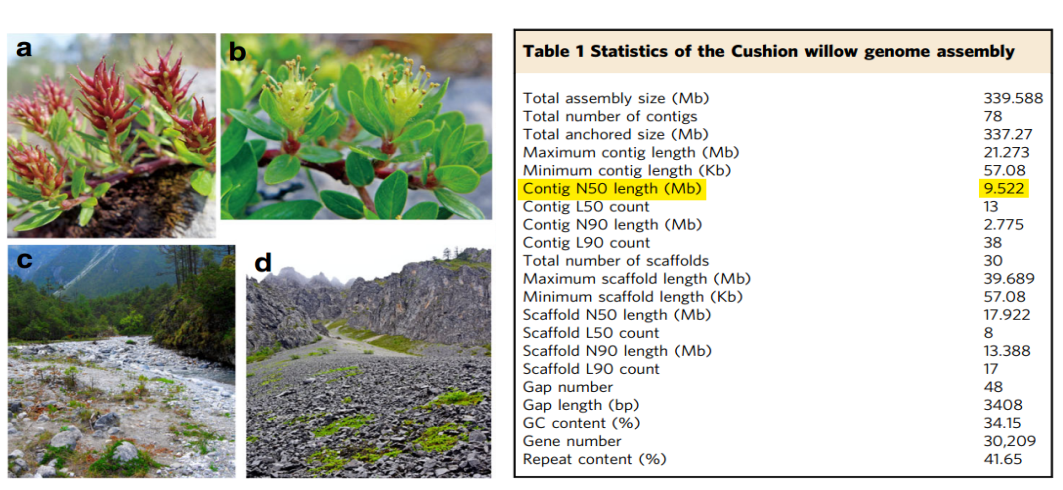
图1 小垫柳基因组组装结果
群体进化研究:群体材料选择(重测序,~22.1x),77个小垫柳(14个主要栖息地);
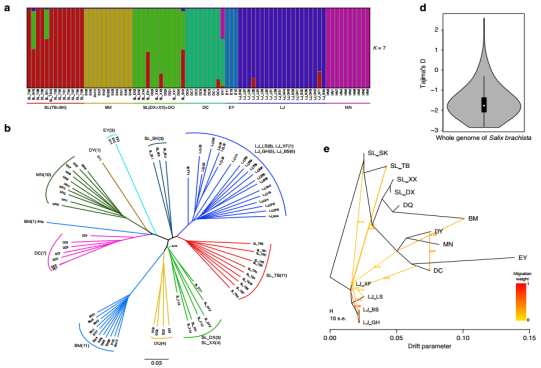
图2小垫柳群体进化分析
研究结果1:小垫柳具有显著的种群分化,尽管其具有长距离传播的能力,但种群间的基因流微弱;小垫柳自晚中新世以来的群体波动与剧烈的气候波动耦合,天空岛效应、多样而异质的生境和气候波动可能是驱动小垫柳种内群体分化的重要因素;
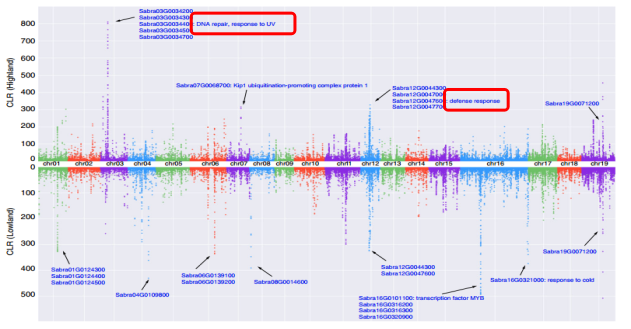
图3小垫柳群体选择(高/低海拔)分析
研究结果2:自然选择在小垫柳高、低海拔群体中的作用区域和强度都具有显著的差异,说明分布于横断山区海拔高差大、生境多样而异质的物种可能在自然选择的作用下发生种下的种群分化(Sabra3G0034400DNA修复,Sabra12G0047600细菌防御应答,Sabra16G0101100细胞对DNA损伤刺激的应答和对寒冷的应答)。
林木基因组重点研究思路(二)
基因组+全基因组关联分析(GWAS)
案例:海枣树基因组

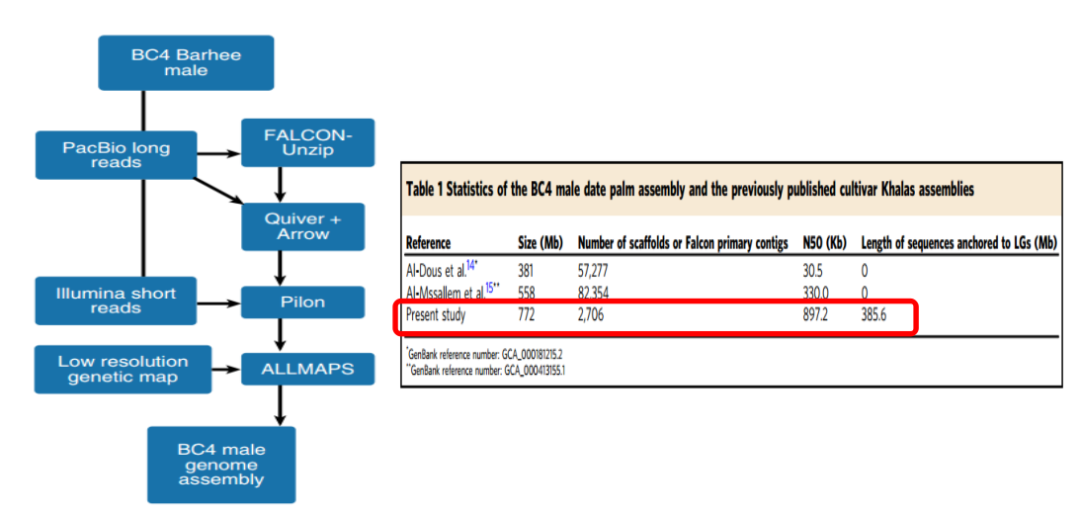
图1 海枣树(BC4雄株)基因组组装
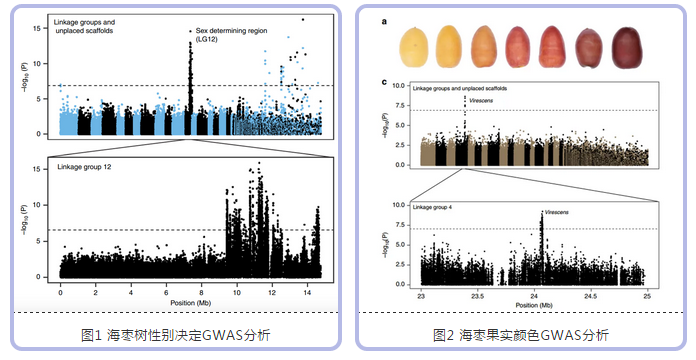
群体GWAS研究:群体材料选择(重测序,~11.3X;):157个海枣品种( 145 个雌性品种和 12 个雄性个体);
林木基因组杂合度高,此外大基因组的针叶树和一些多倍体的树种难以获得高质量的基因组序列,北京百迈客生物科技有限公司拥有丰富的基因组组装经验,尤其对高杂合,大基因组及多倍体植物基因组研究中独具优势,由北京百迈客新推出的ONT+PB CCS的双平台组装策略赢得了众多老师的好评,针对林木基因组组装更具优势,目前公司仅林木类基因组合作文章发表20余篇,累计影响因子达116+,而正是这些高质量林木基因组的破译,将打破林木基因组育种中的瓶颈,为育种研究奠定了深厚的基础。
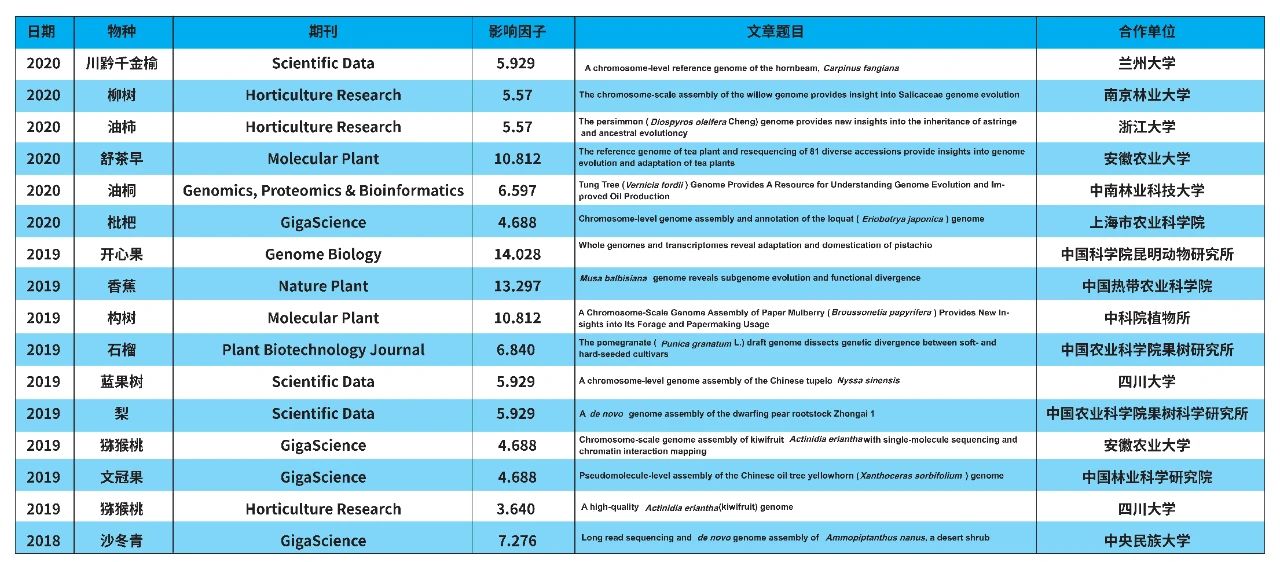
北京百迈客林木研究基因组合作文章(部分)




 京公网安备 11011302003368号
京公网安备 11011302003368号