英文名:Long Non-coding RNA Derived from lncRNA– mRNA Co-expression Networks Modulates the Locust Phase Change
期刊:Genomics Proteomics Bioinformatics
IF:7.051(1区)
通讯作者单位:中科院、河北大学
研究背景
长链非编码RNA(lncRNA)调节基因表达、动物行为等各种生物学过程。尽管蛋白质编码基因、microRNA和神经肽在亚洲飞蝗的表型可塑性调节中起着重要作用,但有关lncRNA在此过程中功能研究较少。本文应用高通量RNA-seq来比较蝗虫型变时程中lncRNA和mRNA的表达模式。结果显示lncRNA在型变的早期阶段反应更快。功能注释表明,早期改变的lncRNA在分散和群居阶段采用了不同的途径来应对种群密度的变化。筛选了分散和群居阶段的网络中两个重叠的中枢lncRNA基因座进行功能验证。本文进一步证明LNC1010057为潜在的蝗虫型变因子。这项工作为深入了解蝗虫型变的分子机制并扩大lncRNA在动物行为中的作用范围提供了重要的数据。
材料方法
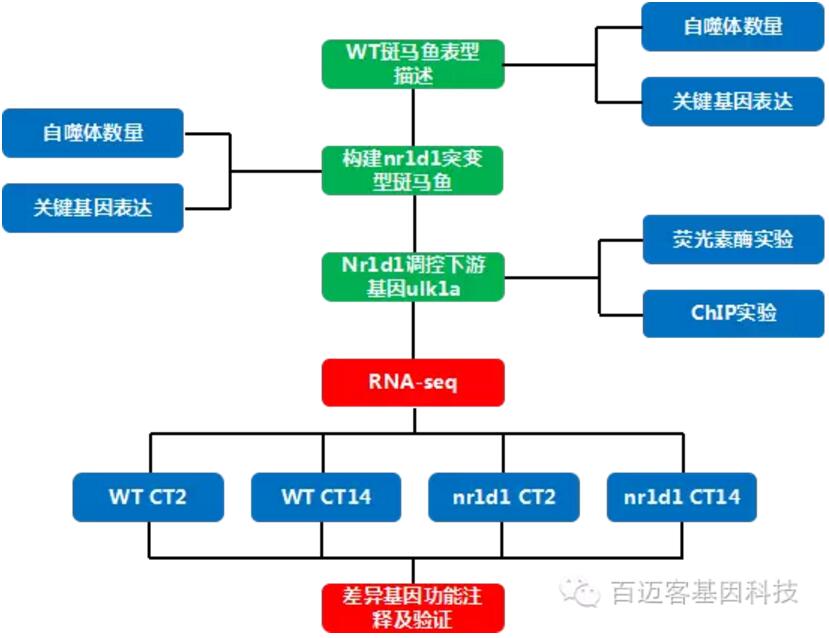
实验材料:散居化处理为将群居型蝗虫单独饲养0h、4h、8h和16h后取出手机蝗虫大脑进行测序;
群居化处理为将10只散居型蝗虫与20只群居型蝗虫饲养于一个小笼子(10厘米×10厘米×10厘米)中,于0h、4h、8h和16h后取出,收集蝗虫大脑进行测序。在同一时间点采集样品的三个生物学重复,提取RNA,建立cDNA文库并进行RNA-seq。对lncRNA和mRNA进行差异表达分析及qRT-PCR验证,对不同时间点的样本进行STEM(ShortTime-seriesExpressionMiner)分析,并构建lncRNA–mRNA共表达网络的构建。对目标中心节点lncRNA进行RNAi,采集其行为录像数据进行行为数据分析。
实验结果
1、蝗虫lncRNA的外显子比mRNA少但更长
研究发现大约78%的lncRNA包含2个外显子,而mRNA中包含的外显子数量为1至120(图1C)。因此,lncRNA的外显子明显长于mRNA的外显子(平均长度1142bpvs.263bp,图1D,左)。同时,lncRNA的内含子明显短于mRNA的内含子(平均长度:10086bpvs.12442bp,图1D,右)。表达水平分析表明,lncRNA的总体表达水平明显低于mRNA的表达水平(平均值为0.6vs.1.9;图1E)。但是,表达特异性分析表明,lncRNA的表达受时间限制的程度要高于mRNA(平均值为0.672对0.436图1F)。基于相对于mRNA的lncRNA基因组位置,蝗虫lncRNA被分类为基因间、重叠区、有义内含子、反义内含子、有义外显子和反义外显子。蝗虫77%以上的lncRNA是长基因间的ncRNA(lincRNA,图1G)。这些结果表明,蝗虫lncRNA与mRNA在结构和表达上有很大不同。蝗虫lncRNAs更长,具有更少但更长的外显子和更短的内含子。此外,lncRNA的表达模式显示出比mRNA更高的时间特异性。

图1lncRNA和mRNA之间的不同结构和表达
2、群居型蝗虫中特异性表达的lncRNA比在散居型蝗虫中表达的更多
在散居型和群居型蝗虫大脑中共表达9722个lncRNA(73.9%),而散居型蝗虫中特异性表达962个lncRNA(7.3%),群居型蝗虫中特定表达2469个lncRNA(18.8%)(图2A,顶部)。分别在散居型和群居型蝗虫中特异性表达了479和1090个mRNA(3.1%和7.1%)(图2A,底部)。这些结果表明,与mRNA相比,在两个蝗虫相型特异性表达的lncRNA的百分比更高,而在群居蝗虫中比散居型蝗虫中表达的lncRNAs更多。与散居型蝗虫相比,群居型蝗虫中335个lncRNA和779个mRNA的表达水平下调,而313个lncRNA和261个mRNA的表达上调[倍数变化(FC)>2和P<0.05;图2B]。lincRNA在下调和上调的lncRNA中所占的比例高(分别为81.2%和87.8%),其次分别是反义外显子和有义内含子lncRNA(图2B)。在表达中按FC排名的前10个lncRNA基因显示在图2C中。这表明散居型和群居型蝗虫大脑中表达的lncRNA的数量及其表达水平明显不同。
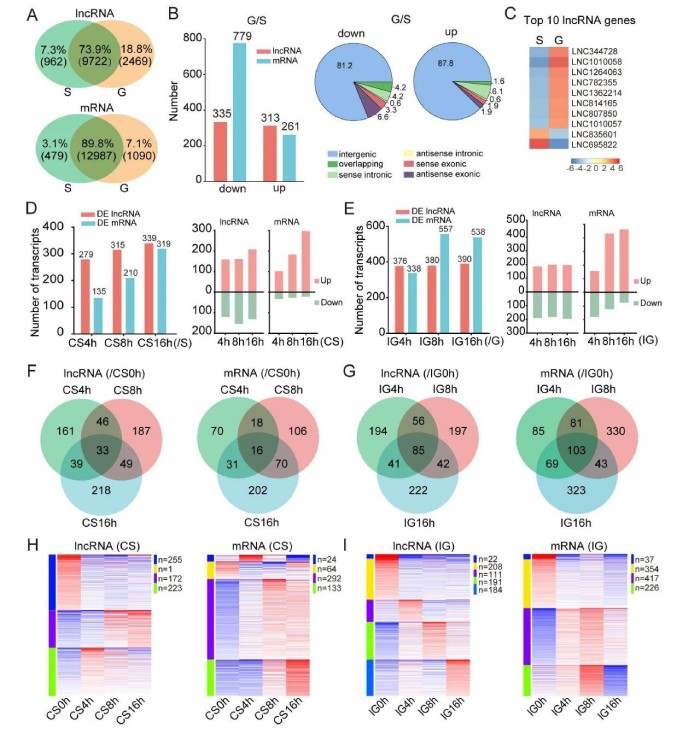
图2lncRNA在蝗虫型变中显示出不同的表达变化模式
3、lncRNA对种群密度变化的快速应答
为了进一步分析lncRNA和mRNA的表达模式,使用了短时间序列表达挖掘器(ShortTime-seriesExpressionMiner,STEM)对基于表达的转录本进行聚类。在CS期间,将214个lncRNA和242个mRNA分别分为8个和9个重要的表达谱(图3D)。其中lncRNA表达谱15以及mRNA表达谱15、13和16直到群居化处理后8或16小时才改变。这些配置文件称为后期更改的配置文件(模式d)。与后期更改的配置文件相反,早期更改的配置文件以4小时时间点开始的转录表达变化为特征。根据4小时后的表达变化,将早期变化的轮廓细分为模式a(早期变化)、模式b(早期中变化)和模式c(可持续变化)。在IG过程中,269个lncRNA和639个mRNA分别聚集成6个和7个显着表达谱。所有lncRNA配置文件均为早期变化(图3E)。在mRNA表达谱中,有6个是早期改变的,而谱12是晚期改变的。在CS和IG期间,lncRNA的聚类分布图的数量与mRNA的分布无明显差异。但是,通过计算谱图中的转录本数量,发现早期改变的lncRNA的百分比比CS中的mRNA的百分比高61.1%(88.3%vs.54.8%,图3F)。同样,IG中早期改变的lncRNA的百分比高于IG中的mRNA(100%vs.81.3%,图3F)。因此,lncRNA比mRNA对散居化和群居化处理的反应更快。在CS中,早期改变的lncRNA的表达变化速率快于285个mRNA的表达变化速率(0.55vs.0.24)和IG(0.77vs.0.39)(图3G)。因此,早期改变的lncRNA的比例和表达变化率高于早期改变的mRNA。因此,蝗虫lncRNA比mRNA对种群密度的变化更敏感。
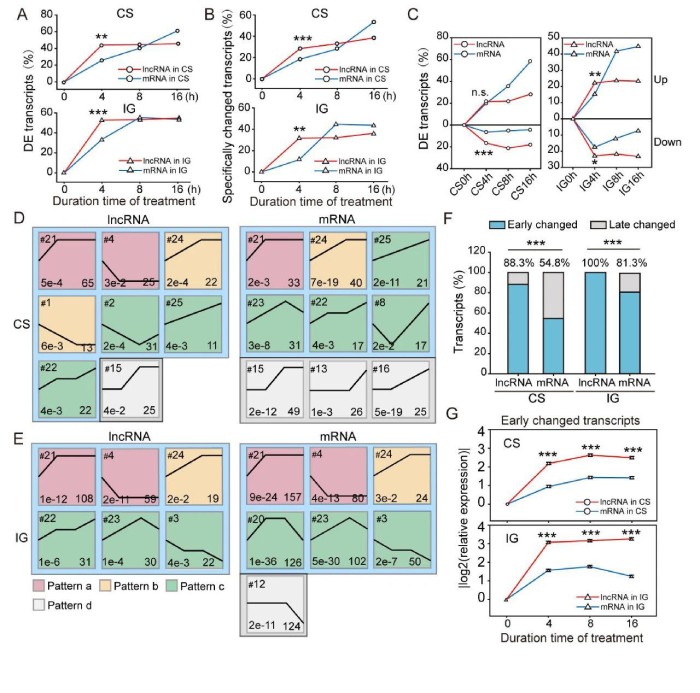
图3lncRNA对散居化和群居化处理的快速反应
4、lncRNA参与CS和IG早期变化的不同途径
在CS中,自噬调节、剪接体和肌醇磷酸代谢途径在上位和至少两个模块中均过代表(图4C)。因此,CS中早期改变的lncRNA可能在调节自噬、RNA剪接和信号转导途径中发挥重要作用。lncRNA对自噬的调控参与了群居化的初期和中期,而对剪接体和磷酸肌醇代谢的调控则参与了整个过程。与神经递质释放有关的突触小泡循环途径的lncRNA仅在模式a(早期改变)模块中被过代表(图4C)。该结果表明,早期改变的lncRNA可能在CS期间调节神经系统中具有重要作用。此外,一些lncRNA与已知的型变相关基因密切相关。例如,早期改变的lncRNALNC531328.2、LNC1425451.2、LNC1088763.7和LNC492755.1与基因NPYR、NPF1a和Vat1相关(图4A)。持续变化的lncRNALNC494161.1和LNC1065048.14与多巴胺途径中的基因Ebony和Vat1相关。在CS网络中,程度值前5%的lncRNA被视为hublncRNA(图4A)。计算了lncRNA基因座的程度值,在CS中,基于度数排名前5%的10个lncRNA基因座被鉴定为hublncRNA。其中,四个基因表达上调,其他六个基因表达下调(图4D)。在IG期间,包括谷氨酸能突触、多巴胺能突触和胆碱能突触途径在内的与突触有关的途径得以丰富。这些途径中的大多数都富含早期改变的模块。同时,多巴胺能突触和胆碱能突触途径仅参与模式a(早期改变)的模块(图4E)。IG中也丰富了信号转导途径,例如钙信号传导、Ras信号传导、胰岛素信号传导和MAPK信号传导途径。功能注释的结果表明,与CS相比,IG中早期变化的lncRNA参与的突触相关和信号处理途径更多。
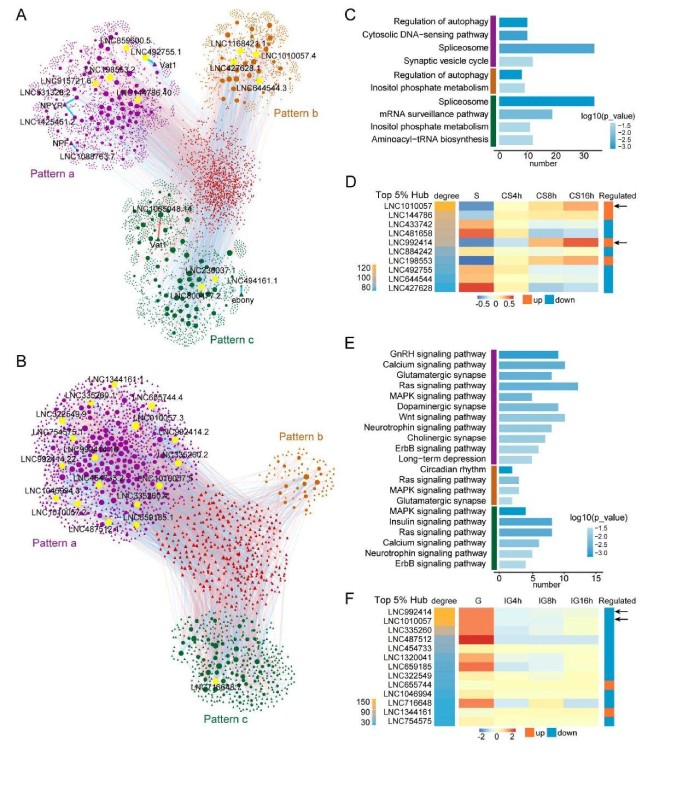
图4早期改变的lncRNA参与CS和IG的不同途径
5、LNC1010057可能调节蝗虫型变
为为了验证LNC1010057和LNC992414在蝗虫型变中的功能,进行了一系列分子生物学实验。首先,克隆了在LNC1010057和LNC992414基因座中鉴定出的长的转录本。LNC1010057由重复的A、B和C元素组成,这些元素依次分布并重复4次,但C元素重复三次(图5A)。序列比对证明LNC992414在5’端与LNC1010057共享相似的重复序列,但是它们是从不同的基因组基因座转录而来的(图5A)。尽管LNC992414的定量表达水平可以通过特异性引物检测,但LNC1010057的表达水平为重复元件的总表达水平。如预期的那样,LNC1010057和LNC992414的表达模式极为相似。在CS期间,两种lncRNA的表达持续增加,并且在16h几乎增加了四倍(图5B)。在IG期间,4h后表达水平显着下降,之后保持相对稳定(图5B)。LNC1010057和LNC992414之间的序列结构和表达模式的相似性表明它们是同源的lncRNA。但是,实时qPCR分析表明,在散居型和群居型蝗虫的大脑中,LNC1010057的表达水平比LNC992414的表达水平高约1000倍(图5C)。其次,为了测试LNC1010057和LNC992414是否参与蝗虫型变调节,在通过RNAi抑制蝗虫大脑中的表达后进行了行为分析。显着降低LNC1010057的表达水平(7470对3764610)和LNC992414(1.9vs.1.0)(图5D)后行为分析表明,群居型蝗虫其行为显着改变为散居状态(图5E)。此外,多个与型有关的行为参数发生了变化。总移动距离(TDM)和总移动持续时间(TDMV)显着降低(图5F),但是,移动速度没有区别。同时群居蝗虫的特定喜好行为显着下降(58.3对-13.5图5F)。但LNC992414表达降低并未引起从群居状态到散居状态的转变(图5G和H)。与对照相比,与型相关的行为参数(包括运动和特定物种的优选行为)没有差异(图5I)。LNC992414的RNAi实验不会引起行为变化,因此排除了LNC992414调节蝗虫型变的可能性。这些结果表明是LNC1010057而不是LNC992414潜在地调节了蝗虫的型变。
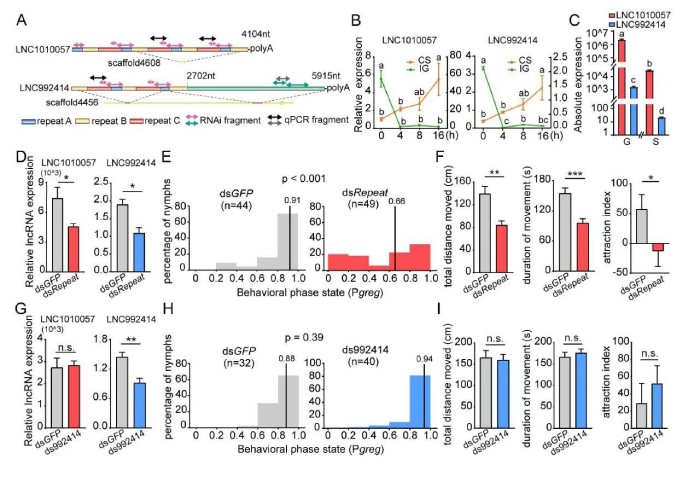
图5LNC1010057潜在地调节了蝗虫的型变
总结
lncRNA被证实是生物过程的关键调控因子,调节包括mRNA转录、稳定性、翻译和翻译后修饰等多种生物途径。之前研究表明昆虫lncRNA参与了例如杀虫剂抗性、繁殖力和腺体凋亡等生物过程,但是尚未有证据证明lncRNA可以调节非模型昆虫的行为。蝗虫是世界范围内的一种农业害虫,表现出显着的表型可塑性。为了鉴定与其型变相关的lncRNA,本文系统地分析了蝗虫lncRNA的表达并注释其功能,证实与mRNAs相比lncRNAs显示对型变更敏感的响应,并证实了其中一个lncRNA可调节型变相关行为。本研究揭示了lncRNA在蝗虫型变中的重要作用以及表型中蛋白编码基因和lncRNA之间的相互作用。深入解析蝗虫散居型和群居型转变的分子机制,为可持续治理蝗虫的新策略和新方法的开发提供了基础。
]]>英文名:The circadian clock regulates autophagy directly through the nuclear hormone receptor Nr1d1/Rev-erbα and indirectly via Cebpb/(C/ebpβ) in zebrafish
杂志:Autophagy,2016
影响因子:9.108
研究背景
细胞自噬是一种非常保守的细胞内降解系统,近期的研究表明小鼠中细胞自噬现象表现出了生物钟节律,nr1d1基因缺陷能导致小鼠骨骼肌中自噬活动的紊乱。然而生物钟调节细胞自噬的机理还不清楚。斑马鱼的细胞自噬活动是否表现出生物钟节律还没有报道,本研究旨在以斑马鱼为实验材料研究nr1d1基因在调节细胞自噬的生物钟节律方面的作用。

实验设计
实验材料:2种材料:野生型WT和nr1d1突变型斑马鱼
2个时间点:CT2和CT14(分别为受精后122和134小时);
无生物学重复
测序方法:百迈客Illumina HiSeq2000 PE125
结果展示
1.表型描述及关键基因在幼鱼和肝脏中的表达情况
首先用TEM(透射电镜)对一天内不同时间(ZT0,ZT6,ZT12,ZT18)的肝脏切片进行观察,统计一天内自噬体数量的变化(红色箭头处,图1,A),并进行统计(图1,B),研究表明斑马鱼肝脏内自噬体的数量符合生物钟节律。
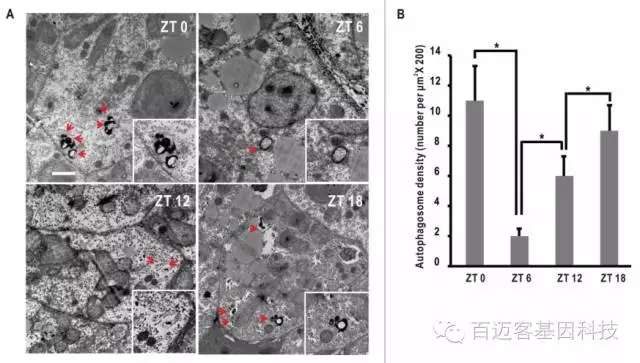
TEM观测及自噬体数量统计结果
选取了几个以前研究过的与自噬体形成、移动、延伸、融合、降解有关的基因进行RT-PCR分析,如maplc3b,bnip3,becn1等。材料为受精后72-96小时的斑马鱼幼体和肝脏材料。由图可见这些基因都表现出明显的生物钟节律。图中白色底色表示白天,灰色底色表示黑夜。
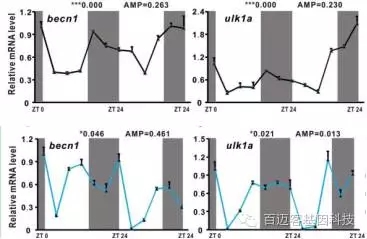
图2 关键基因的RT-PCR检测
2.用TALEN技术构建nrid1突变斑马鱼,并进行表型研究和关键基因表达分析
用TEM观察WT和nr1d1突变斑马鱼肝脏中的自噬体数量,研究表明自噬体数量在nr1d1中明显高于WT。
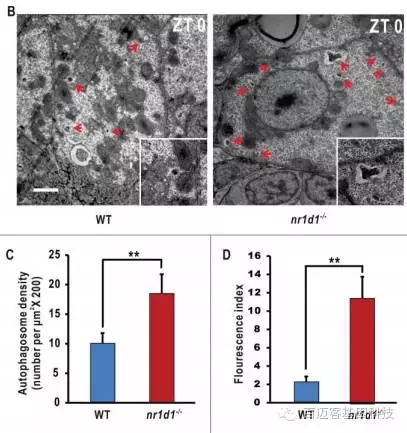
图3 不同样品斑马鱼肝脏自噬体观测及统计
用RT-PCR研究在WT和nr1d1突变斑马鱼中关键基因的表达差异,nr1d1突变体中基因表达模式与WT中明显不同。
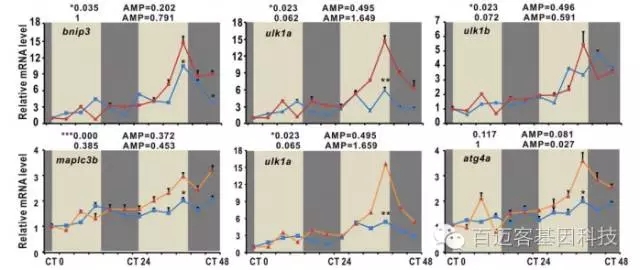
图4 差异基因的RT-PCR验证
3.用ChIP和荧光素酶研究Nr1d1直接调控的下游基因
体外荧光素酶实验表明Nr1d1蛋白能直接结合ulk1a基因的promoter区域,体内ChIP实验进一步证明Nr1d1蛋白能够结合ulk1a基因的promoter区域。说明Nr1d1能直接调控ulk1a基因的表达。

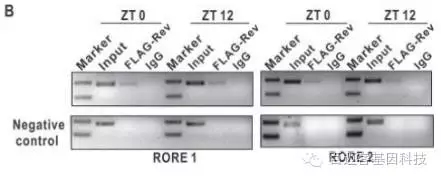
图5 荧光素和chip结果
4. 野生型WT和nr1d1突变型斑马鱼转录组分析
为了进一步说明nr1d1基因的功能,文章选取了野生型WT和nr1d1突变型斑马鱼进行转录组研究,并且取了2个时间点(CT2和CT14)。研究表明在CT2阶段两者有975个差异表达基因,在CT14阶段有1360个差异表达基因。与WT相比,在nr1d1突变体中上调的基因数量要大于下调的基因数量。
同时,选取部分关键的差异表达基因进行RT-PCR验证,结果与转录组分析结果一致。
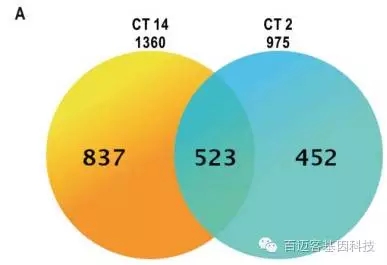
图6 差异基因韦恩图

图7 差异基因聚类图
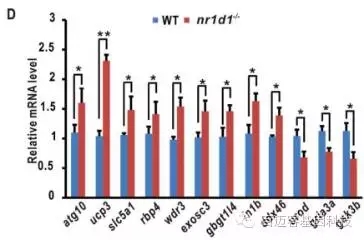
图8 关键基因的RT-PCR验证
研究结论
通过研究WT野生型和nr1d1突变型斑马鱼的基因表达情况,表明Nr1d1基因在生物钟调节和细胞自噬过程中起着重要作用。生物钟相关基因直接受Nr1d1调控。此外,通过转录组分析表明Nr1d1还调控胁迫、代谢、信号传导、翻译后修饰等生理过程。
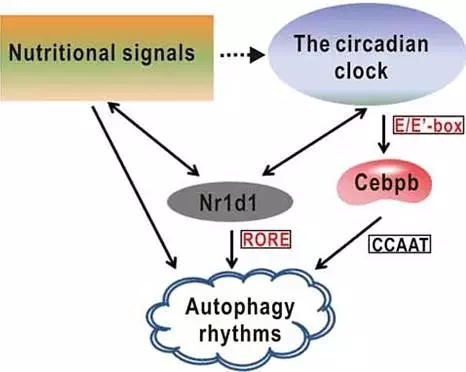
图9 Nr1d1在生物钟和细胞自噬过程中的作用模式图
亮点
1.首次利用斑马鱼研究生物钟和细胞自噬的关系;
2.发现Nr1d1在生物钟调节和细胞自噬过程中的关键作用;
3.转录组研究进一步证明Nr1d1的作用,并揭示Nr1d1其它生理功能。
4.虽然转录组的内容在这篇文章中比重不大,但却揭示了除了生物钟调节和细胞自噬,Nr1d1基因还有很多其它的功能,显著提升了文章的水平,并为后续研究打好了基础。
英文名:Transcriptome analysis of Chlorella zofingiensis to identify genes and their expressions involved in astaxanthin and triacylglycerol biosynthesis
杂志:Algal Research
影响因子:5.014
研究背景
小球藻为单细胞可食用绿藻,能够合成虾青素和油脂;虾青素是一种结构独特的酮式类胡萝卜素,是自然界已知最强的抗氧化剂。含油脂高的藻类可作为生物质能源。
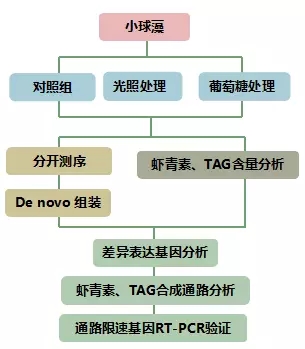
实验设计
材料选择:小球藻 取种子细胞培养到生长对数期的后期,分成3组,用于做不同处理;
处理方式:对照组T1:不做处理;
高光处理组T2:光照90μmol光子?2 s?1,6h;
葡萄糖诱导组T3:30g/L葡萄糖处理, 6h;
测序方法:Hiseq2500,4G/样;
技术路线:
研究结果
1. 虾青素和脂类定量分析
对照和葡萄糖处理组相比,葡萄糖诱导下的小球藻中,虾青素含量96h后增加了30倍,而处理组中96h后只增加了8倍。脂类和TFA含量也有类似的情况。对照组中这三种成分增加较少可能是培养基中营养成分限制的原因,在氮饥饿的情况下,导致在相同器官中虾青素和脂肪酸产量增加。
2. RNA-SEQ分析
三种不同的处理构建三个文库,测序获得clean data 10.63G,Trinty组装获得32931个unigenes。52.8%的unigenes在NR,Swiss-Prot,KEGG,GO,COG获得注释。
3. 差异表达基因分析
计算RPKM,做差异表达分析。T1与T2差异表达基因数为580,T1与T2差异表达基因数为1489。在光处理和葡萄糖处理两种情况下,差异基因功能注释表明这些基因都是与翻译、核糖体、蛋白过程、胁迫和碳固定相关的。在光处理下,卟啉和叶绿素代谢、光刺激应答是被富集的。在葡萄糖处理情况想,葡萄糖、能量产生和转化、脂肪合成等都是被富集的;而这些基因在光合作用、核糖体、蛋白运输通路中是减少的。在碳源充足的情况下,生长和能量存储基因是上调表达的,而光合作用是下调表达的。
4. RT-PCR验证
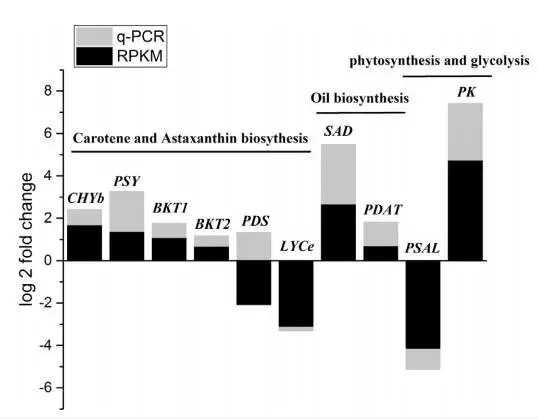
作者选了10个与虾青素和三酰甘油合成有关的unigenes进行了RT-PCR验证,其中9个基因的表达趋势与RNA-seq结果一致。只有PDS两种结果不同,可能是RNA-seq结果不准确,因为前人的研究成果中,葡萄糖是上调PDS表达的,与本文中RT-PCR结果一致。
5. 虾青素合成
在转录组测序结果中,发现MEP通路中的8个基因都被鉴定到,但是在MVA通路中只有2个基因被检测到,其中HMG表达不显著,ACAT表达量下降。这些结果表明小球藻中虾青素的合成通路中IPP和DMAPP是MEP通路合成的。其中,发现一个可能编码β-胡萝卜素激酶(BKT2)与数据库中的BKT1比对率仅为56%。功能分析发现BKT2可以转化β-胡萝卜素成为角黄素。下图为虾青素合成通路图。在葡萄糖处理情况下,与虾青素合成相关的6个基因上调差异表达。在小球藻中,葡萄糖诱导虾青素的合成,伴随着BKT1,CHYb,PDS在24h内上调表达。为研究转录组水平基因表达量与产物含量的关系,分析了5个限速基因的表达量,这些基因在葡萄糖处理72h或92h表达量达到最高,相应的,虾青素的积累量也在96h达到稳定水平。
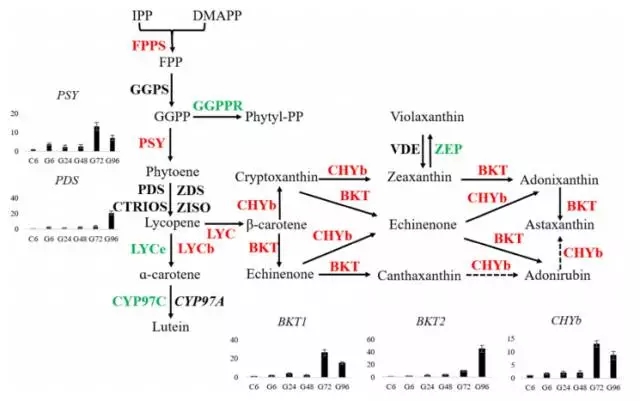
6. 三酰甘油的合成
藻类和植物中脂肪酸的生物合成主要在叶绿体中进行。乙酰-CoA作为合成的起始物质。基于转录组测序,小球藻质体中脂肪酸合成基因被鉴定到。在葡萄糖处理情况下,这些基因大都表达量上调。另外这次测序结果中还发现几个基因对应的不同转录本。由于BC和SAD是脂肪酸合成的限速酶基因,做了进一步分析,发现BC和SAD表达量从6h到72h表达量持续上升,对应的三酰甘油含量从24h到96h持续增加。
三酰甘油(TAG)合成有两条通路:乙酰-CoA依赖的通路和乙酰-CoA不依赖的通路;转录组数据中两条通路中的基因都被鉴定到。
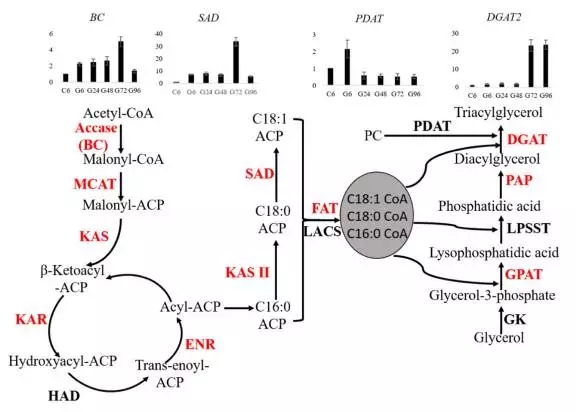
文章亮点
1:材料选择+处理条件;
小球藻关注的成分具有重要的经济价值,处理条件是可以促进经济成分积累,研究更加有利用价值;
2:实验数据+前人结果,深入验证;
测序结果中重点关注关键成分(虾青素和TAG)合成通路中的每一个基因,对其进行差异分析,并做不同时期的表达量分析;解释清楚在处理情况下,物质合成与基因表达的关系。针对本文的研究结果与前人研究成果比较分析。
]]>
在瘤胃中的短链脂肪酸(SCFAs)对反刍动物的生长和健康起着关键的作用,微生物G蛋白偶联受体(GPR)和微生物脱乙酰化酶(HDAC)可能是这些影响的主要调节通路。该研究主要是选用不同比例非纤维碳水化合物摄入(15-30%)的山羊模型,通过转录组测序和16srRNA测序联合研究瘤胃上皮细胞和瘤胃微生物协同的响应机制。通过研究发现,瘤胃中微生物来源的SCFAs对上皮细胞的生长和代谢受到GPR和HDAC调节网络的影响,通过对这些调控机制和饮食组成的相互关系的理解,可更深入的了解瘤胃的新陈代谢机制,更好的促进牧业的发展。
样品取材:共6只雄性山羊,随机分为两组,第一组饲养为65%的干草+35%的饲料(MC组),另一组为90%的干草+10%的饲料(LC组);喂养28d后进行取样。
微生物多样性样品:饲养第28天,在清晨喂养0h、2h、5h以及8h进行取样,瘤胃内容物用4层纱布取样 ,收集瘤胃液15ml,-20℃保存用于提取。
转录调控测序取样:取10cm2瘤胃组织,用冷却的PBS进行清洗,去除肌肉层,取上皮组织切块于TRIzol保存液保存。液氮速冻5min后,-80保存用于RNA提取。
短链脂肪酸(SCFAs)浓度测定:上述样品加入5%HgCl2用于浓度测定。
测序分析:
瘤胃内容物微生物多样性测序:细菌V3+V4区、Illumina Miseq 平台测序
瘤胃上皮细胞转录组测序测序: PE125 测序、Illumina Hiseq2500 平台测序
1、短链脂肪酸(SCFA)浓度变化分析:
饲养不同处理组(MC和LC)、喂养取样不同时间点间瘤胃内总短链脂肪酸(SCFA)、PH以及短链脂肪酸(SCFA)主要成分的变化情况进行分析,如下图。

?图1.1?总SCFA变化情况
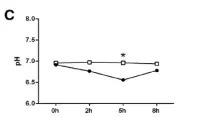
图1.2?PH变化情况
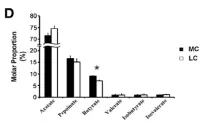
图1.3?SCFA主成分变化情况
2、瘤胃微生物多样性分析:
2.1细菌群落结构分析
在细菌门水平,一共有14个原核的门被鉴定,主要是厚壁菌门(35.7-30.1%),拟杆菌门(26.6-43.6%)、互养菌门(11-7%),且通过门水平维恩图分析发现,在两组(MC和LC)间门水平组成相同,比较分析两组间微生物变化情况发现(如图:FIG2),MC组相比于LC组,互养菌门增长了57%,无壁菌门增长了330%。而黏胶球形菌门降低了63%,纤维杆菌门降低了65%;

图2.1?两组间门水平Venn图分析
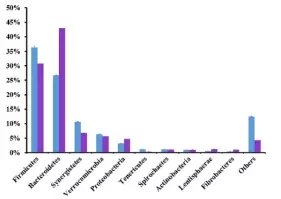
图2.2 两组间门水平OTU分析
在细菌属水平。共鉴定出75个属水平细菌,在两组间共有70个属,在MC组有三个特有的属,在LC水平有2个特有的属,普氏菌属(10.4-17.9%)在两组间都为主要属水平的菌。
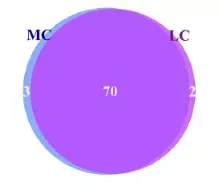
图3.1 两组间属水平Venn图分析
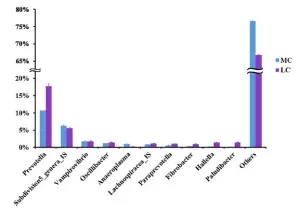
图3.2?两组间属水平OTU分析
2.2 微生物群落的多样性和丰富性
在门水平,MC组的拟杆菌门和厚壁菌门显著性高于Lc组(P<0.05),互养菌门显著性低于Lc组(如下图4),通过最大似然法(ML)计算27个丰度大于1% 的OTU进行分析显示,在MC组显著增加的OTU属于子单胞菌科,疣微菌科、韦荣球菌科、疣微菌门,相反的,58%(7/12)显著降低的OTU属于普雷沃氏菌科(如下图5);
?图4
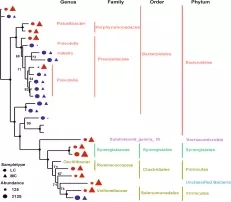
图5
3.瘤胃表达谱分析:
3.1 瘤胃上皮细胞GPRs和HDACs的表达谱分析
RNA-Seq测序方法用于研究山羊瘤胃微生物G蛋白偶联受体(GPRs)和微生物脱乙酰化酶(HDACs)表达情况以及调控网络,过滤得到127M clean data,平均有83%数据比对到NCBI山羊参考基因组,得到73个GPR家族成员和11个HDAC家族成员比对到山羊基因组,转录组数据分析发现有20个GPR家族成员和7个HDAC家族成员在瘤胃上皮细胞中表达。通过比较LC组中基因的表达发现GPR1,87,89A,155显著上调(P<0.05),在MC组中GPR107,游离脂肪酸受体4(FFAR4, also known as GPR120),羟基烃酸受体2(HCAR2, also known as GPR109A)显著上调(P<0.05),通过组内基因表达分析,在两组内都发现GPR家族中GPR87表达*高,HDAC家族中HDCA1的表达*高
3.2 GPRs和HDACs保守序列分析
为了研究GPR和HDAC在进化过程中差异表达的保守序列,通过用所有基因进行ML进化树分析,所有的 GPRS和HDACS在脊椎动物门都是高度保守的。研究GPR家族内成员的保守序列发现,在GPR家族成员没有超过30%的相似性,在HDAC家族成员之间也发现了同样的结果。
图6
3.3 差异表达的GPRS和HDACs基因相关KEGG通路分析
为了研究GPR和HDAC共表达网络的生物学意义,改研究注释了相关的信号通路和KEGG功能,发现显著差异的邻近基因被分类到瘤胃上皮细胞的生理调节过程,因此发现了两类和上皮细胞生长调节有关系的功能网络,其中一类是上皮细胞生长网络(包括细胞凋亡,增殖和分化,见下图7),另外一类是上皮细胞的代谢网络(包括辅酶因子和维生素代谢,能量代谢、氨基酸代谢等,见下图8);
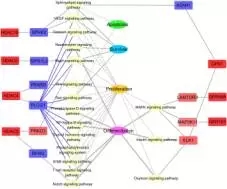
图7:上皮细胞生长网络图
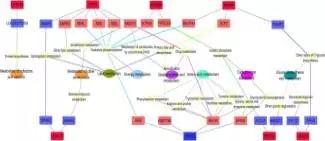
图8:上皮细胞代谢网络图
在上皮细胞生长网络中,基因GPR1,89和155的表达上调导致LAMTOR3蛋白,蛋白激酶和ELK1表达上调,这三种蛋白都存在于MAPK信号通路上,和细胞的增殖分化有关。此外,GPR1的 表达增加和酸性酰胺酶(ASAH1)的下调相关,ASAH1存在于调节细胞凋亡、生存和增殖的信号通路上。对于HDACs家族基因HDAC4,5,6和10基因的上调和五个调节细胞凋亡,生存和增殖基因的下调有关。
在上皮细胞代谢网络中,GPR1、87和89A基因的上调表达与辅酶2、4以及4L(ND2,4,4L),ATP5H,NDUFA4有关,HDAC1的下调表达ND6的上调表达有关。以上所有相关的酶都和能量代谢的氧化磷酸化有关。
4.转录调控和16s rRNA数据联合分析?
GPR和HDAC的表达、SCFAS主成分浓度比例以及属水平细菌相对丰度关系分析
通过CCA相关性分析发现,显著上调的GPRs和HDACs与8个显著增加的微生物属(56个 OTUS)是呈正相关,显著下调的GPRs和HDACs和9个显著减少的细菌属呈正相关(63个OTU),然而HDAC1与丁酸盐的比例呈负相关。
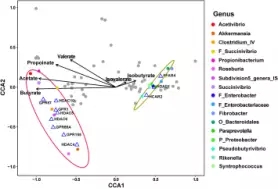
该研究得到的结果揭示,SCFA调节的GPR和HDAC共调控网络存在于瘤胃的上皮细胞,该共调节网络作用可作用于动物敏感√确地接受微生物区的信号调节,这些调控网络调节上皮细胞各种生理学过程,尤其是细胞的生长和新陈代谢,在促进动物的生长和维持上皮细胞的完整性起到关键的作用。此外,这些调控网络重要的调控机制对于共生的细菌来说,主要通过调节上皮细胞生理过程提高细菌在宿主中的共生条件。通过了解宿主动物和微生物区之间互相的调节机制,加上相应饮食中的外界干预调节因素,可以可持续性的更好的提高动物的健康和生长。
Hong Shen, Zhongyan Lu, Zhihui Xu, Zhan Chen,nd Zanming Shen Associations among dietary non-fiber carbohydrate, ruminal microbiota and epithelium G-protein-coupled receptor, and histone deacetylase regulations in goats.Microbiome.2017;5:123