英文名:Phylogenetic tree-informed microRNAome analysis uncovers conserved and lineage-specific miRNAs in Camellia during floral organ development
杂志:Journal of Experimental Botany,2016
影响因子:IF=5.526
研究背景
在植物中miRNA是源自有发夹结构单链前体的内源性小RNA。miRNA和其靶基因的进化代表了一种动态的基因表达通路,它们在决定植物不同器官发育中起基础性的作用。本文利用高通量小RNA测序的手段研究了山茶花5个器官的miRNA空间表达谱。
实验设计
材料:Camellia
取样组织:五个组织材料(幼嫩叶片、雄蕊、花瓣、心皮和花芽),三个生物学重复
测序策略:百迈客Illumina HiSeq? 2500 ,SE36
测序数据量:每个样本测了18.9~28.2M的数据量
设计思路:
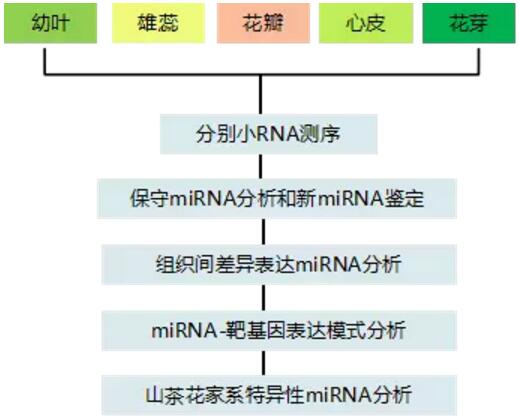
研究结果
1、山茶花5个组织保守miRNA分析和新miRNA鉴定
miRDeep2软件鉴定175个潜在的miRNA。长度分布分析显示21和22bp的miRNA丰度*高;对成熟miRNA的碱基偏好性分析:21和22bp的miRNA首位碱基偏好于“U”;23和24bp的miRNA首位碱基偏好于“A”。
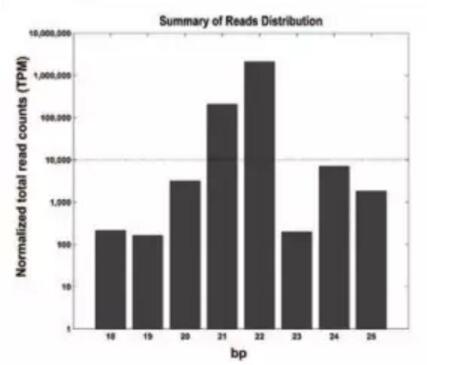
miRNA长度分布
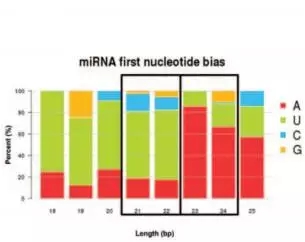
miRNA碱基偏好性
2、山茶花中MIR160家族的结构和进化分析
将成熟的miRNA比对到植物的ncRNA Database鉴定到19个miRNA。以MIR160家族为例进行系统进化分析:基于pre-miRNA序列来构建MIR160家族系统进化树。结果显示保守的c38436.graph_c0_79977与来自葡萄、杨树、木薯和莲花的MIR160形成了一个分枝,这也与山茶花的进化地位相一致。
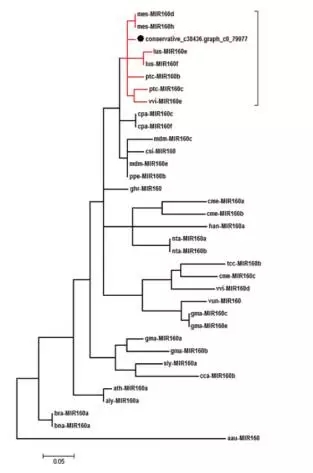
miRNA 160系统进化树
3、组织间miRNA差异表达分析

组间差异表达miRNA维恩图
4、组织特异性miRNA鉴定
对不同组织miRNA表达情况绘制聚类热图,大部分miRNA在不同组织中特异表达。miRNA表达聚类热图显示一些明显有着不同表达模式的亚簇,在此主要关注了雄蕊和心皮所特异的2个亚簇。对雄蕊中特异表达的miRNA靶基因进行GO注释,显著富集GO term为“DNA intergration ”和“氧化还原”。
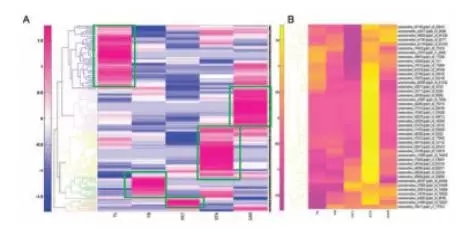
miRNA表达量聚类热图及GO注释
5、miRNA-靶基因的表达相关性验证
选取6个miRNA及其靶基因进行表达验证,发现4对表现出负相关的关系。例如miRNA167靶向2个SPL基因,用qPCR进行验证,结果显示出强的负相关关系。

miRNA及靶基因表达量相关性
6、山茶花中家系特异的miRNAs
(1).为了鉴定山茶花中家系特异miRNA家族,对不同植物中的miRNA家族构建进化树。发现2个葡萄特异的miRNA家族出现在山茶花中,其中miR3633是高丰度和高识别度的,表明它出现在葡萄和山茶花分化之前。
 miRNA家族进化树
miRNA家族进化树
(2).鉴定到12个山茶花家系特异miRNA,将其前体序列与NCBI中山茶花和葡萄转录组数据库进行比对。在葡萄数据集里没有发现相应的miRNAs,但是大部分都出现在其他山茶花种里,说明这12个miRNAs是以家系特异的方式进化的。
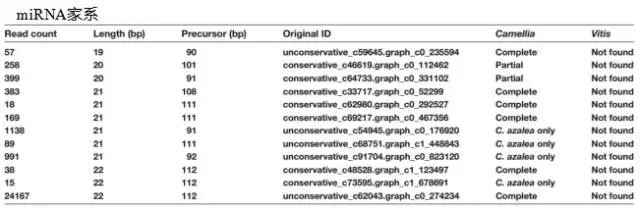
文章亮点
1:材料选择;
山茶花是观赏植物,因其在不同类型重瓣上产生额外的花瓣形成它独特的观赏价值。了解其花器官发育的分子机制有助于新品种的培育。
2:深度数据挖掘,实验验证;
从组织特异性和发育调控方面,鉴定了组织特异性保守miRNA,深入进行靶基因功能注释富集分析揭示其参与器官形成的机制;重点关注控制花器官发育的miRNA-靶基因调控机制,并选择一些关键miRNA-靶基因进行了实验验证,为miRNA-靶基因环状调控方式提供了依据。
3:新的分析角度;
从进化角度对山茶花属的miRNA进行了分析,描述了山茶花属miRNA家系特异型的进化方式。
Shoot bending promotes flower bud formation by miRNA-mediated regulation in apple (Malus domestica Borkh.)
Plant Biotechnology Journal,2015
实验结果
开花诱导在苹果树生命周期中发挥非常重要的作用,但是年轻的苹果树往往容易产生低质量的数量少的花芽。嫩枝弯曲可以促进产生较多的花芽,因此嫩枝弯曲成为苹果树一个非常重要的栽培性状。树嫩枝弯曲会产生一种新的长距离信号,从而改变树的生长和发育。但是,响应嫩枝弯曲发生的花芽生长和开花诱导的分子调控机制并不清楚,尤其是miRNA是否在其调控中发挥作用以及具体的机制如何。本研究关注在花芽发育、开花诱导和发育、响应嫩枝弯曲过程中miRNA的潜在作用。
研究目的
解析苹果树响应嫩枝弯曲时,植物激素和miRNA协同调控花芽生长和开花诱导的分子机制,有助于解决年轻苹果树低质量花芽的问题。
材料方法
1.实验材料:
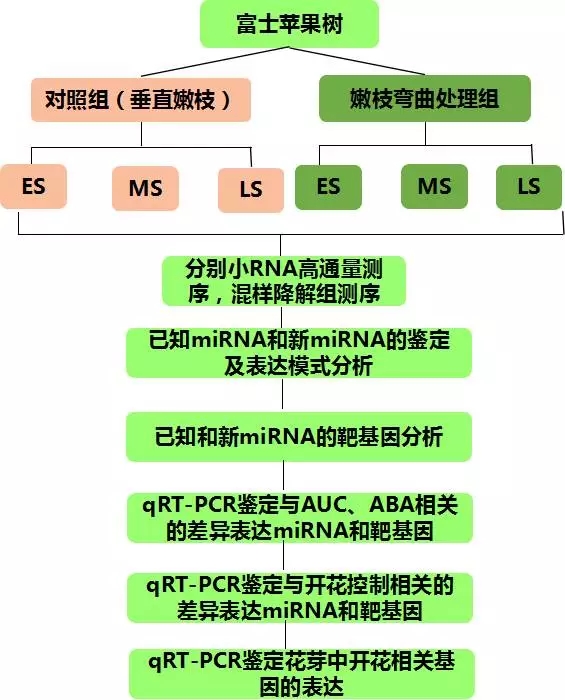
生长六年大小的富士苹果树,分两组进行嫩枝弯曲(110度)、对照处理(垂直嫩枝),分别在开花诱导的初期(early stage,ES)、中期(Middle stage,MS)和晚期(Later stage,LS)取花芽,分别提取RNA进行小RNA测序,6个花芽组织RNA混样进行降解组测序。
2.测序方法:
小RNA seq,Biomarker Illunima Hiseq 2000 platform
技术路线
实验结果
1. 富士苹果树在开花诱导时期响应嫩枝弯曲后的花芽生长
首先,研究富士苹果树在开花诱导时期,嫩枝弯曲处理后花芽生长情况。无论是对照还是嫩枝弯曲处理,从发育早期(ES)到晚期(LS),花芽的长度、宽度、干重都增加了。但是,嫩枝弯曲明显增加了花芽的大小。此外,通过检测花芽生长速率发现,花芽大小变化主要发生在花芽诱导的早期,即ES到MS时期;而且,在嫩枝弯曲处理下花芽生长速率明显增加了。实验结果还显示,嫩枝处理组比对照组的开花速率明显增大了。
2. 开花诱导时期花芽中激素含量的变化。
分别在开花诱导的三个时间点(ES, MS, LS),对嫩枝弯曲处理组和对照组的花芽中激素(生长素,AUX;脱落酸,ABA;细胞分裂素,CK;赤霉素,GA)含量进行检测。分析了在嫩枝弯曲组合对照组,开花诱导过程中各种激素含量的变化。
3. 小RNA和降解组文库构建和测序。
为了研究嫩枝处理和对照组在ES、MS和LS时期小RNA的情况,构建了6个miRNA文库(CES, CMS, CLS, BES, BMS, BLS)并用Illunima Hiseq 2000进行了测序。总共获得了14 095 388, 14 106 904, 15 242 433, 13 259 311, 15 969 504 和13 523 103 raw reads。此外,对这六个组织提取的总RNA构建了降解组文库并进行了测序,总共获得了30 762 927 (93.08%) clean reads。分别对文库进行了插入片段大小分布的评估,对sRNA的长度进行了统计分析。
4. 已知miRNA的鉴定和表达模式分析。
为了鉴定苹果中已知的miRNA,用BLASTN将这六个花芽的sRNAs比对到miRBase 18.0 和植物miRNA数据库中。总共获得了属于41个miRNA家族的195个已知的miRNA。然后对已知miRNA的表达量进行了分析。

Fig1. 差异表达已知miRNA维恩图
对同一处理不同发育时期样品间(CMS vs CES, CLS vs CMS, CLS vs CES, BMS vs BES, BLS vs BMS and BLS vs BES)筛选了差异表达miRNA并绘制了维恩图。结果显示,其中42个miRNA下调表达,20个miRNA上调表达。对不同处理相同发育时期样品间(BES vs CES, BMS vs CMS and BLS vs CLS)筛选到124个差异表达miRNA,相对对照组,嫩枝弯曲处理组中68个下调表达,27个上调表达。
5. 新miRNA的鉴定和表达模式分析。
用栽培种苹果(Malus domestica Borkh)作为参考基因组来预测潜在的新miRNA,共鉴定到137个新miRNA。对同一处理不同发育时期样品间(CMS vs CES, CLS vs CMS, CLS vs CES, BMS vs BES, BLS vs BMS and BLS vs BES)新miRNA进行差异表达分析并绘制了维恩图。结果显示,其中34个新miRNA随着发育时期上调表达,24个新miRNA在嫩枝弯曲处理组表达高于对照组,23个新miRNA在对照组表达明显高于嫩枝弯曲处理组。
对嫩枝弯曲处理组和对照组间的31个差异表达新miRNA进行表达层次聚类分析。结果显示,根据表达模式可分为5个亚簇。其中Cluster 1中的6个新miRNA在嫩枝弯曲组表达量均高于对照组。Cluster 5中的新miRNA表达模式则与Cluster 1中相反。其他三簇中的19个新miRNA,无论在对照组还是嫩枝弯曲处理组,展现出较低的表达水平。
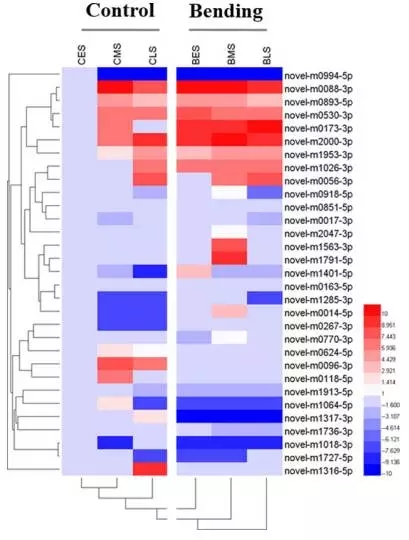
Fig2. 差异表达新miRNA聚类热图
6. 已知和新miRNA的靶基因分析。
为了研究鉴定的已知和新miRNA参与的生物学过程,以及分析嫩枝弯曲调控开花诱导的分子机制,通过降解组来鉴定miRNA的靶基因。上文的分析显示,在对照组和嫩枝弯曲组之间筛选到了41个家族195个已知miRNA。对这些已知miRNA以及它们目标切割位点的具体分布信息进行了分析。同时,鉴定了31个差异表达新miRNA的40个靶标。
新miRNA调控的靶基因包括一些转录因子和调控蛋白。如,新m1736-3p的靶基因为编码一个myb-like的HTH转录因子家族蛋白(DUO1)。一些新miRNA调控的靶基因与植物激素相关,如新m1791-5p调控的靶基因为参与CK信号转导途径的细胞分裂素响应因子CRF4。此外,一些新miRNA的靶基因参与糖信号转导和代谢,如新miR1316-5p调控靶基因BGAL3。
7. 定量RT-PCR鉴定与AUC、ABA、控制开花相关的差异表达miRNA和靶基因
为了验证miRNA测序结果的准确性,以及分析在不同发育时期(ES-MS-LS)miRNA和靶基因的表达水平,我们检测了在响应嫩枝弯曲时与IAA、ABA、控制开花相关的miRNA和靶基因的表达模式。结果显示,与IAA、ABA信号转导相关的miRNA和靶基因可能参与调控响应嫩枝弯曲时苹果花芽的形成。
创新点
1.苹果树嫩枝弯曲可以促进产生较多的花芽,嫩枝弯曲是苹果树一个非常重要的栽培性状,研究解析苹果树响应嫩枝弯曲的分子机制,有助于解决年轻苹果树低质量花芽的问题。
2.结合降解组的数据来分析鉴定miRNA的靶基因,更具有说服力。
3.分析了嫩枝弯曲诱导开花过程中激素含量的变化,并集合miRNA测序数据针对性地分析了激素相关基因的调控作用,深入解析了分子机制。
4.大量qRT-PCR实验对miRNA和降解组测序数据的验证。
参考文献
[1] Libo Xing, Dong Zhang, Caiping Zhao, Youmei Li, Juanjuan Ma, Na An and Mingyu Han, (2015) Shoot bending promotes flower bud formation by miRNA-mediated regulation in apple (Malus domestica Borkh.) Plant Biotechnology Journal, pp. 749–770.
]]>实验材料
大豆“Taiwan 75”培养在正常条件下,第一真叶期的幼苗分为对照组和处理组。对照组—25°C(12h),17°C(12h),处理组—4°C(24h)。设置3次生物学重复。
研究结果
1.sRNA测序数据统计
首先,对大豆4°C处理组(3h、6h、9h、12h、24h和36h)及对照组进行相对生长速率(RGR)和MDA含量的检测。发现在24h时,对照组和处理组RGR和MDA含量明显不同。因此,选择冷胁迫24h进行sRNA测序。
对照组和处理组分别获得6559098和8273245 clean reads。长度为21nt所占比例*高(图1A)。非冗余的reads,长度分布统计可以看出24nt所占比例*高,在对照组和处理组中分别为68.4%和59%(图1B)。
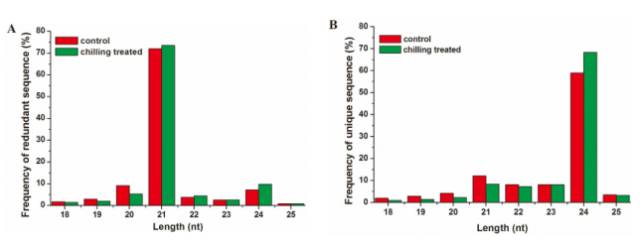
图1.sRNA长度分布图
2.鉴定大豆中已知的miRNA
共鉴定到已知的434个miRNA,属于133个家族。这些miRNA包括保守miRNA和种间特异性miRNA。保守miRNA在植物发育过程和响应胁迫上起到重要作用。通过同源比对鉴定大豆中保守的miRNA家族。例如miR156,miR160,miR164等在很多植物物种中都是高度保守的。此外,找到一些非保守miRNA,例如miR3522,表明他们可能参与大豆的物种进化。
3.预测大豆中新的miRNA
根据miRNAs前体的发夹结构来预测miRNAs,找到3个预测的miRNA并鉴定折叠成的二级结构。
4.验证大豆中预测的miRNA
为了验证测序结果,利用qRT-PCR分析miRNA表达情况。选择了35个miRNA(33个已知miRNA,2个预测miRNA)进行qRT-PCR分析。线性回归分析测序结果和qRT-PCR结果相关性系数为0.8048,表明miRNA-Seq和qRT-PCR结果相一致。尽管miRNA-Seq和qRT-PCR结果在√确的倍数方面有些差异,这可能是由于2种实验结果敏感性和特异性的不同导致的,但是miRNA表达趋势是一致的。
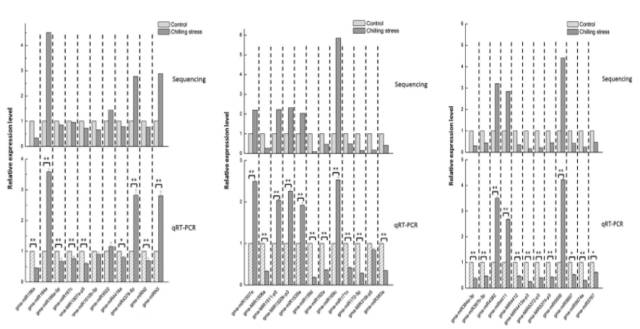
图2.qRT-PCR结果
5.miRNA靶基因鉴定
利用降解组测序鉴定miRNA降解的靶基因。共获得12283683条raw reads,2623291条非冗余raw reads。共鉴定到898个转录本是54个miRNA家族的靶基因。本研究发现miRNAs可以降解2个甚至更多的靶基因,与之前的研究相似。例如,gma-miRNAs可以沉默属于SBP家族的15个基因,脱落酸响应结合因素蛋白家族和2个基因,2个转录因子等。而且,有7个转录本受到多个miRNAs的调控。
6.鉴定大豆冷胁迫下相关的miRNA
对miRNA进行差异表达分析,共鉴定到32个miRNA家族的51个miRNA差异表达。在冷胁迫下有30个下调表达,21个上调表达。
对相应靶基因进行功能注释来研究差异表达的miRNA可能行使的功能。差异表达的miRNA可以分为四类。第一类包括miRNA家族(miR156、miR164、miR169、miR4412和miR5327),靶基因为转录因子SBP、NAC、NFY、GRAS和bHLH,参与调控基因表达和信号传导。第二类包括miR4411,其对应靶基因涉及抵御疾病。第三类包括miR5761、miR159、miR5667、miR1535、miR511、miR4382和miR4416,其对应靶基因参与植物生长发育中的适应性。为了进一步确认冷胁迫下miRNA和靶基因的关系,选择了5个miRNAs及其靶基因进行qRT-PCR验证。结果表明,在冷胁迫下miRNA-164a,miRNA-4411和miRNA-169e上调。相反的,他们的靶基因NAC、DRP和NFY显著下调。说明miRNA和对应靶基因呈负相关性。
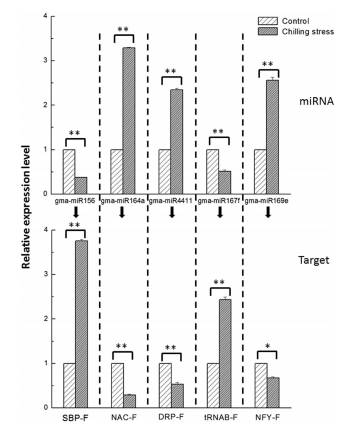
图3.miRNA和对应靶基因qRT-PCR图
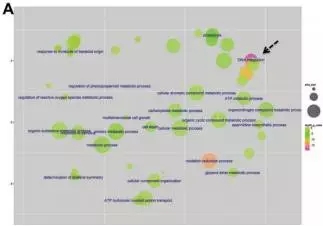
为了近一步确认这些差异表达miRNA的功能,对靶基因进行GO分析。靶基因共参与了56个分子功能terms,37个生物学过程terms和7个细胞组分terms。在分子功能分类下,ATP结合、蛋白结合、组蛋白结合等terms是显著富集的。超过35%miRNAs的靶基因参与了生物学过程途径。

图4.GO注释
结 论
首次研究大豆冷胁迫下miRNA调控基因表达情况。利用降解组测序鉴定到了上百个靶基因,揭示miRNAs和靶基因之前的相互关系。尽管miRNAs调控机制十分复杂,目前还不是十分清楚,本次研究完善了miRNA数据库,为研究大豆和其他物种的基因调控网络提供了重要基础。也发现了51个响应冷胁迫的miRNAs。
文章亮点
1.处理时间点的选择上进行了预实验,设置多个时间梯度进行处理并进行生理指标测定,最终选择变化较为明显的时期。选样时间点有理有据。
2.利用降解组测序z确鉴定miRNA降解的mRNA。
参考文献
[1] Xu S, Liu N, Mao W, et al. Identification of chilling-responsive microRNAs and their targets in vegetable soybean (Glycine max L.)[J]. Scientific Reports, 2016, 6.